Prairiesoilsandcrops.ca

Prairie Soils &
Insects and Diseases
Crops Journal
Managing the Pea Leaf Weevil in Field Peas
Héctor Cárcamo* and Meghan Vankosky
H. A. Cárcamo, Agriculture and Agri-Food Canada, Lethbridge, AB; M. Vankosky, University of Windsor, Windsor, ON
*Corresponding author E-Mail:
Summary
The pea leaf weevil is one of the few insect pests of field peas in southern Alberta and southwest Saskatchewan.
Adults cause distinctive notching on seedlings which generally does not affect yield. Larval feeding on nitrogen-fixing
root nodules may reduce yield if soil is nitrogen deficient. Only field peas and faba beans are at risk of damage. Other
crops in the bean family may be fed on by adults but do not suffer significant damage from larvae. The economic
threshold during the second to fifth node stages in field peas is 30% of seedlings with damage to the terminal (clam)
leaf. Seed treated with thiamethoxam has produced more consistent yield responses than application of foliar
insecticides but additional farm-level trials are required. Trap cropping the pea leaf weevil can be done by planting a
border in the fall with winter peas or adjacent, earlier planting of a spring cultivar. Close monitoring of the trap crop is
needed to control the weevils in the trap crop area before they move to the rest of the field. Fields under no tillage
regimes suffer less damage than those under conventional tillage. Biological control is a research area that should be
pursued for sustainable management of this pest.
Background
The pea leaf weevil, Sitona lineatus L. (Fig. 1), belongs to the most speciose insect order, the Coleoptera, and the most
speciose beetle family, the Curculionidae. Although over 100 Sitona species occur worldwide, only 11 are found in
North America and five of these were introduced1. The pea leaf weevil is a common pest of field peas, Pisum sativum
L. and faba beans, Vicia faba L. (Fabales: Fabaceae) in its native Europe2. It was first collected on Vancouver Island in
19363. Since then it has been reported in several states on both the east and west coast of the USA1,4, southern Alberta
in 1997 (R. J. Byers, personal communication) and Saskatchewan in 20075. A detailed review of its history, biology
and management has been described by Vankosky et al.6. In this review we focus on its local biology, summarize
research on chemical and cultural management in the southern Canadian prairies and provide a short summary of key
recent publications.
Figure 1: Adult pea leaf weevil showing alternating light and dark brown stripes extending from the thorax to the abdomen (H.
Goulet, AAFC-Ottawa).
Volume 4 ▪ 2011
www.prairiesoilsandcrops.ca

Prairie Soils &
Insects and Diseases
Crops Journal
Morphology and Seasonal Biology
The morphology of all stages of the pea leaf weevil was described in detail by Jackson7. Further morphological details
and development time of each stage in Europe and North America are summarized by Hoebeke and Wheeler8. In the
Canadian prairies, pre-overwintering adults can be distinguished from related Sitona, such as the sweet clover weevil,
by light and dark brown alternating stripes extending along the pronotum onto the base of the elytra (Fig. 1). Older
adults which have lost the scales that form the stripes require examination under a microscope to confirm that the
anterior procoxal cavities touch the narrow groove in front1.
The following summary is based upon field and laboratory studies conducted in the Lethbridge area from 2007 to
20099. Adults were collected in pea fields using pitfall traps or pan traps as early as mid-May in warm years such as
2007 and 2 to 3 wk later in cool years such as 2008 and 2009. Adults immediately fed on pea foliage (Fig. 2), mated
and started laying eggs within a week. Peak egg densities occurred 2 to 3 wk after arrival and ranged from about 2000-
20,000 eggs per square meter. Under laboratory conditions (16/20°C), eggs took 12 to 14 d to hatch. Larval densities
peaked from late June to early July at 2000- 5000 larvae per m2 and declined each year by mid July. Pupal densities
peaked from mid July to early August. Adults lay eggs over 3 months so all stages can be found simultaneously even
though there is only one generation per year. New generation adults start emerging from the soil in late July, with peak
emergence occurring in mid-August. Emergence continues until the host plant senesces. New generation adults do not
mate but do feed on any green legume foliage before seeking sheltered leaf litter habitats for overwintering.
Figure 2: Damage to pea foliage by pea leaf weevil (Photo credit: H. Goulet, AAFC-Ottawa).
78 www.prairiesoilsandcrops.ca Volume 4·▪ 2011

Prairie Soils &
Insects and Diseases
Crops Journal
Host Damage and Nitrogen
Although pea leaf weevil adults have several legume hosts, reproductive potential and larval fitness are maximized on
faba bean and field pea. An adult weevil consumes about 6 mm2 of pea seedling foliage per day10 (Landon et al.,
1995). Feeding results in a characteristic scalloping on the leaf margins (Fig. 2)7,11. Under high infestations at the
early seedling stage, feeding damage can severely reduce stand density7. However, plants can usually compensate for
this damage so yield losses are minimal12. Larvae feed on root nodules that contain Rhizobium leguminosarum bacteria
that fix atmospheric nitrogen (Fig. 3)13.
Figure 3: Larva of pea leaf weevil on a Rhizobium root nodule of field peas (C. Herle, AAFC-Lethbridge).
The impact of pea leaf weevil on nitrogen fixation and uptake by field pea accounts for its pest status. Under controlled conditions, up to 98% of nodules were excavated14. In plots near Lethbridge in 2007 under extremely high weevil pressure (>100 notches/plant), over 90% of the nodules were damaged regardless of the crop stage when an insecticide was applied (Fig. 4). Depending on soil nitrogen levels, larval feeding had a greater impact on yield than adult feeding15,16. In faba beans, yield loss was due to fewer pods rather than fewer or lighter seeds per pod.
Figure 4: Proportion of field pea nodules destroyed by pea leaf weevil larvae near Lethbridge in early- and late-seeded plots in
Volume 4 ▪ 2011
www.prairiesoilsandcrops.ca
Prairie Soils &
Insects and Diseases
Crops Journal
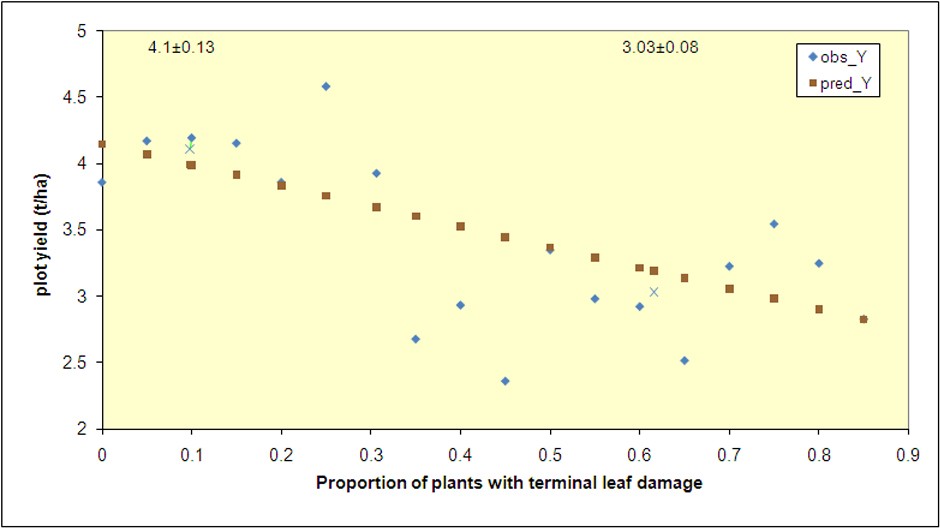
Field plot and cage studies were conducted near Lethbridge and Vauxhall from 2007 to 2009 to validate economic thresholds and assess pea leaf weevil - field pea interactions with soil nitrogen9,17. El-Lafi18 observed significant yield reductions when field pea stands had more than 30% of seedlings with terminal leaf damage. This threshold was validated in Alberta using field pea plots planted with or without thiamethoxam insecticide seed treatment. Plots that had less than 30% seedling damage before the 5th node stage, yielded at least 155 kg/ha more than those with 40% or more damage (Figure 5). Removing the insecticide-treated plots from the analysis did not affect results.
Figure 5: Observed mean pea yield in relation to proportion of plants with damage on terminal leaf (0.05 intervals). Predicted line
from regression with plots with more than 1.2 t/ha or less than 6.9t/ha. Y = 4.145 – 1.553 (x). R2 = 0.16, DF = 1, 140, p = 0.000. "X" symbol denotes average for all plots under or over economic threshold of 0.301, values and standard errors given above Xs.
Mean yields difference between 0.30 and 0.40 damaged plots represents 155 kg/ha. There are no significant differences in yield
among plots with proportion of seedlings with terminal leaf damage between 5 – 30 % (ANOVA, p>0.05) or those with damage
between 30 – 85% (ANOVA, p>0.05).
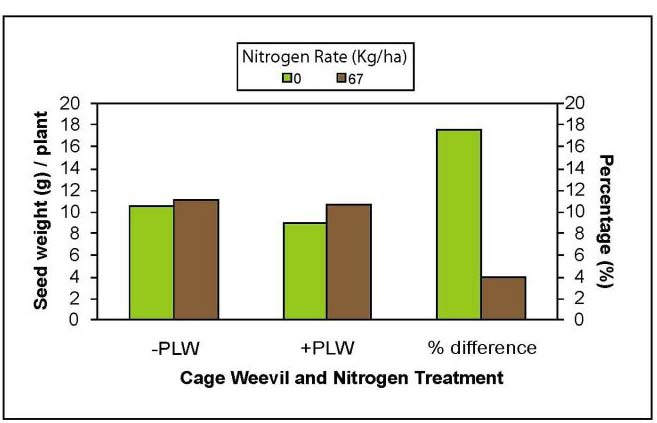
Interactions with nitrogen were more subtle and no significant effects were observed in the field19. However, at the Lethbridge cage study site, plants growing in nitrogen-depleted soil experienced a greater yield loss (17%) than plants that were amended with urea (3% yield loss) (Fig. 6). A greenhouse study to further investigate the effects of larval feeding on plant and soil nitrogen also suggested that soil amendment with urea can buffer the negative effects of S. lineatus on plant nitrogen content and soil nitrogen (Cárcamo, Herle, and Lupwayi., unpublished data). Therefore, if soil nitrogen is sufficient, the pea leaf weevil poses less of a risk and control measures are not required.
80 www.prairiesoilsandcrops.ca Volume 4·▪ 2011
Prairie Soils &
Insects and Diseases
Crops Journal
Figure 6: Effect of soil amendment with urea at 67 kg/ha on the ability of caged pea plants to tolerate feeding damage by pea leaf
weevil. Entries are averages of cages 1 m2 with 32 plants with 0.25. 0.50, or 1 pea leaf weevil (+PLW treatment, densities pooled)
per plant or zero weevils (-PLW treatment).
Monitoring and Chemical Management
Adult pea leaf weevils are difficult to monitor because they drop to the ground when approached and their cryptic color
makes them difficult to see against the soil. Despite these challenges a nominal threshold of 0.3 to 1 adult weevils per
seedling has been recommended20. To count adults, the observer should sit quietly and still for a short time until
weevils resume moving. During the warmest hours of the day, usually late morning to mid-afternoon, weevils are
easily observed walking from plant to plant over the soil surface (Cárcamo, field observation). Species confirmation
requires adult collection but threshold determination can be easily assessed by counting seedlings with terminal leaf
feeding. At least 5 locations along the field margin and 5 locations approximately 100 m into the field should be
selected to assess pea leaf weevil damage for threshold determination. At each location, a row of 10-20 seedlings
should be selected and the terminal leaf on each seedling examined. If the average proportion of seedlings with
terminal leaf damage at all spots exceeds 30% (i.e., more than 3 of 10 seedlings, or 6 out of 20), then the weevil poses
a yield risk and warrants control. Field peas should be monitored from the second to fifth node growth stages. After
the sixth node, plants are less vulnerable to both larval damage as shown in a greenhouse study (Cárcamo and Herle,
unpublished data) and defoliation in a field study12. An aggregation pheromone has been used to monitor dispersal of
pea leaf weevil, but trapped numbers could not be related to yield losses21. The pheromone monitoring system could
be used to detect adult weevils in regions where it is expected to invade.
Managing the pea leaf weevil after damage reaches the economic thresholds is a challenge. Several foliar insecticides
were tested in southern Alberta (Meers, unpublished data) from 2007 to 2010 but no yield benefits were observed. The
same result was observed in plots near Lethbridge (Fig. 4) where a pyrethroid insecticide was applied at various crop
stages to early- and late-seeded field peas9. Historically, products including permethrin, cypermethrin, carbosulfan,
imidacloprid and cyhalothrin lambda have been tested against pea leaf weevil, with variable results when applied to the
foliage (reviewed in 6). An insecticide with a longer residual period is likely to be more effective given the protracted
spring migration and invasion time of this pest over several weeks in the spring22. However, these products also pose a
high risk to the environment and non-target natural enemies, hence, most have been de-registered.
Volume 4 ▪ 2011
www.prairiesoilsandcrops.ca
Prairie Soils &
Insects and Diseases
Crops Journal
Insecticides applied in-furrow at planting or seed coatings are generally more efficacious than foliar treatments6,9
(Meers, unpublished data). Carbamates such as furathiocarb reduced larval damage and increased yield relative to
untreated plots in Europe23,24, but have been deregistered due to environmental concerns6. Imidacloprid, a
neonicotinoid applied as a seed coating, reduced adult feeding by 50%23. In Alberta, another neonicotinoid seed
treatment Cruiser® (thiamethoxam) was registered on an emergency basis and later received full registration against pea
leaf weevil based on reduced foliar feeding by adults. A series of controlled studies9 showed that thiamethoxam killed
less than 30% of adults. Weevils that appeared to be dead recovered and resumed feeding within a few days, and foliar
damage was reduced by about 50%. Furthermore thiamethoxam delayed oviposition by about one week, reduced
oviposition during the critical crop stages (second to fifth node) by about 90% and killed approximately half of the
larvae feeding on the nodules of treated plants. Field plot studies to assess yield responses to thiamethoxam, however,
have produced inconsistent results19 (Meers, unpublished data). As noted for other seed treatments, thiamethoxam may
be effective at moderate weevil densities but further research at the commercial scale is needed.
In summary, to manage pea leaf weevil in the Canadian prairies, farmers should determine their relative risk from the
distribution map (available at:
ation
on weevil abundance. In regions of high risk, producers may consider using a seed treatment (consult provincial crop
protection guide for latest registered products) when soil nitrogen levels are low. Otherwise, growers can monitor their
fields and if damage exceeds the economic threshold, they may apply a foliar insecticide between the third and fifth
nodes to reduce weevil populations. If spring conditions are warmer than average, control action should be taken
earlier, but if conditions are cool, control may not be needed as weevils will emerge and infest plants that are more
advanced. If infestation occurs after the sixth node, plants are less vulnerable to foliar damage12 or larval feeding on
root nodules.
Alternative Agronomic Management Strategies
Alternative pea leaf weevil management strategies, suitable for use in an Integrated Pest Management program, have
also been studied on the Canadian prairies. In Europe, trap crops have been investigated as a management strategy25.
In Alberta, a two-year trap crop study was conducted near Lethbridge using winter peas as the trap crop (K. Coles,
unpublished data, summarized in 9). Austrian winter peas (Fig. 7) were planted in autumn around the perimeter of
fields that were seeded with spring pea (main crop) the following spring. In 2007, weevil damage to pea seedlings was
very high in the trap crop and was negligible in the spring peas before mid May. Delays in insecticide application to
the trap crop, or extended weevil migration, led to significant damage in the main crop and the entire field had to be
sprayed. Less damage was observed in winter pea (Whistler variety) trap crops in 2008, as was damage in the main
crop. This was probably because cool, wet spring conditions delayed weevil emergence. These are only preliminary
results; however, trap cropping has potential to manage the pea leaf weevil and merits further investigation.
82 www.prairiesoilsandcrops.ca Volume 4·▪ 2011

Prairie Soils &
Insects and Diseases
Crops Journal
Figure 7: Trap cropping pea leaf weevil, near Nobleford in southern Alberta, using Austrian winter peas along the perimeter and
spring peas planted in the rest of the field. Photo credit: Héctor A. Cárcamo, AAFC, Lethbridge.
In addition to trap cropping, the impact of tillage regime on pea leaf weevil has recently been investigated in the United
States. Evidence from several research projects indicate that migrating pea leaf weevil are more attracted to
conventionally tilled plots relative to no-till plots and feeding damage by pea leaf weevil was significantly greater in
conventional till than in no-till plots26,27,28. In conventional till plots, larvae developed faster and new generation adult
emergence occurred earlier than in no-till plots, where larval development was delayed and larval survival to the adult
stage was reduced27. The impact of tillage regime on the yield of field pea attacked by the pea leaf weevil has yet to be
investigated. In addition to reducing pea leaf weevil density, no-till is expected to positively impact the natural enemies
of the pea leaf weevil such as carabid beetles29, which would add to the value of this cultural control tactic.
Biological Control
Biological control of pea leaf weevil has been more thoroughly investigated in Europe than in North America; a
number of potential natural enemies have been identified (reviewed by6). Aeschlimann2 reported that the parasitoid,
Microctonus aethiopoides (Loan) (Hymenoptera: Braconidae) was an important natural enemy of Sitona weevils in the
Mediterranean, and this species is being considered for the control of S. lepidus Gyllenhal (Coleoptera: Curculionidae)
in New Zealand30. A similar parasitoid, M. aethiops Nees and two other braconid wasps, Perilitus rutilus (Nees) and
Pygostolus falcatus (Nees) (Hymenoptera: Braconidae) and the fly, Campogaster exigua (Diptera: Tachinidae) were
released in an attempt to control S. cylindricollis (Fahr), the sweet clover weevil, in Manitoba in the 1950s, but
apparently did not become established31. The only other stage vulnerable to parasitoid attack is the egg stage. The
mymarid wasp, Patasson lameerei Debauche may be a Sitona specialist32 and was responsible for 95% of all attacks of
eggs observed throughout southern Europe but the overall mortality from egg parasitism was generally low and rarely
exceeded 20% in France2. Pea leaf weevil larvae are sheltered in root nodules, but may be preyed upon by several
nematode species with variable effects on mortality33,6. Generalists predators such as ground beetles (Coleoptera:
Carabidae) attack adult weevils, especially as they emerge from the soil34. Small carabid beetles in the genus
Bembidion (e.g., B. quadrimaculatum L.) are well known predators of insect eggs and this species was a voracious
predator of pea leaf weevil eggs under laboratory conditions35. Bembidion quadrimaculatum and other small ground
beetle species are common
Volume 4 ▪ 2011
www.prairiesoilsandcrops.ca
Prairie Soils &
Insects and Diseases
Crops Journal
in agroecosystems36,35. To better promote carabid beetle populations in field pea crops, the application of foliar
insecticides should be avoided as much as possible. Alternatively, systemic insecticides employed as seed treatments
are more compatible with biological control programs37,38,39, although further investigation regarding the efficacy of
systemic insecticides against pea leaf weevil is required.
Conclusions
The pea leaf weevil is well established in the southern prairies but it is unclear if it can expand northward where peas
are grown extensively. Damage to field pea is more likely to occur in early planted fields if spring conditions are
warmer than average and weevil arrival coincides with the vulnerable crop stages. A key area of research for the pea
leaf weevil is population forecasting to assess damage risk at the local level. Currently, risk maps indicate the general
region of potential damage based on spring counts of plant damage. However, it is difficult to determine if a seed
treatment will be cost-effective. At this point Growers need to rely on both the regional map and their local
observations of weevil abundance at the end of the summer. A quantitative predictive model to estimate weevil
populations in the spring and the need to plant seed treated with an insecticide, will require more research on
overwintering survivorship and local sampling of populations at the end of the summer that could be highly laborious.
Over the long term, a biological control program may provide sustainable management if exotic parasitoids of the egg
and the adult stage could be introduced from the native range of this insect.
References
1.
Bright, D. E. 1994. Revision of the Genus Sitona (Coleoptera: Curculionidae) of North America. Ann. Entomol. Soc. Am.
Aeschlimann, J.-P. 1980. The Sitona (Col.: Curculionidae) species occurring on Medicago and their natural enemies in the
Mediterranean region. Entomophaga 25:139-153.
Downes, W. 1938. The occurrence of Sitona lineatus in British Columbia. Can. Entomol. 70:22.
Bloem, S., R. F. Mizell, and C. W. O'Brien. 2002. Old traps for new weevils: new records for curculionids (Coleoptera:
Curculionidae), brentids (Coleoptera: Brentidae) and anthribids (Coleoptera: Anthribidae) from Jefferson Co., Florida.
Fla. Entomol. 85:632-644.
Hartley, S. 2007. Saskatchewan insect pest report. Minutes of the 2007 Western Committee on Crop Pests, Penticton,
British Columbia, 18 Oct. 2007. Onlin 25 Nov.
Vankosky, M., L. M. Dosdall, and H. A. Cárcamo. 2009. Distribution, biology and integrated management of the pea leaf
Sitona lineatus L. (Coleoptera: Curculionidae), with an analysis of research needs. CAB Reviews: Perspectives in
Agriculture, Veterinary Science, Nutrition and Natural Resources. doi: 10.1079/PAVSNNR20094007.
Jackson, D. J. 1920. Bionomics of weevils of the genus Sitones injurious to leguminous crops in Britain. Ann. Biol. 7:269-
Hoebeke, E. R., and A. G. Wheeler, Jr. 1985. Sitona lineatus (L.), The pea leaf weevil: first records in eastern North
America (Coleoptera: Curculinoidae). Proc. Entomol. Soc. Wash. 87:216-220.
Cárcamo, H., M. Vankosky, C. Herle, S. Meers, K. Coles, R. McKenzie, and O. Olfert. 2010. Integrated pest management
of the pea leaf weevil in field peas: Final report for ACIDF, Alberta and Saskatchewan Pulse Growers Commission and
Landon F, J. Levieux, J. Huignard, D. Rougon and P. Taupin. 1995. Feeding activity of Sitona lineatus L. (Col.,
Pisum sativum L. (Leguminosae) during its imaginal life. J. Appl. Entomol. 119: 515-522.
Nielsen, B. S. 1990. Yield responses of Vicia faba in relation to infestation levels of Sitona lineatus L. (Col.
Curculionidae). J. Appl. Entomol. 110:398-407.
Williams, L., D. J. Schotzko, and L. E. O'Keeffe. 1995. Pea leaf weevil herbivory on pea seedlings: effects on growth
response and yield. Entomol. Exp. Appl. 76:255-269.
Johnson, M. P., and L. E. O'Keeffe. 1981. Presence and possible assimilation of Rhizobium leguminosarum in the gut of
pea leaf weevil, Sitona lineatus, larvae. Entomol. Exp. Appl. 29:103-108.
El-Dessouki, S.A. 1971. The effect of some Sitona spp. larvae on leguminous plants. Z. Angew. Entomol. 67:411-431.
Doré, T., and J. M. Meynard. 1995. On-farm analysis of attacks by the pea weevil (Sitona lineatus L.; Col. Curculionidae) and the resulting damage to pea (Pisum sativum L.) crops. J. Appl. Entomol. 119:49-54.
84 www.prairiesoilsandcrops.ca Volume 4·▪ 2011
Prairie Soils &
Insects and Diseases
Crops Journal
Y.05. N2 fixation and N supply in organic pea (Pisum sativum L.) cropping systems as
affected by weeds and peaweevil (Sitona lineatus L.). Eur. J. Agron. 22:449-458.
Vankosky, M. A. 2010. Integrated pest management of Sitona lineatus L. (Coleoptera: Curculionidae) in crops of Pisum
sativum L. (Fabales: Fabaceae) in western Canada. Masters Thesis, University of Alberta, Edmonton, Alberta, Canada. 18.
El-Lafi, A. M. 1977. The economic injury level of the pea leaf weevil, Sitona lineatus (L.) in northern Idaho and eastern
Washington. Ph.D. dissertation, University of Idaho, Moscow.
Vankosky, M. A., H. A. Cárcamo, R. H. McKenzie, and L. M. Dosdall. 2011. Integrated management of Sitona lineatus
L. (Coleoptera: Curculionidae) with nitrogen fertilizer, Rhizobium inoculation and thiamethoxam insecticide. Agron. J. (in
Antonelli, A., A. Retan, L. E. O'Keeffe, and C. Johansen. 1985. Pea leaf weevil: its biology and control. Extension
Bulletin, Washington State University [0903], 3 pp.
Nielsen, B. S., and T. S. Jensen. 1993. Spring dispersal of Sitona lineatus: the use of aggregation pheromone traps for
monitoring. Entomol. Exp. Appl. 66:21-30.
Hamon, N., R. Bardner, L. Allen-Williams, and J. B. Lee. 1987. Flight periodicity and infestation size of Sitona lineatus.
Ann. Appl. Biol. 111:271-284.
Steene, F., G. Vulsteke, M. de Proft, and D. Callewaert. 1999. Seed coating to control the pea leaf weevil, Sitona lineatus
(L.) in pea crops. Z. Pflanzenk. Pflanzen. 106:633-637.
Taupin, P. 1994. Protection insecticide – La précision génère la marge. Perspectives Agricoles 187:3 pp.
Smart, R. R., M. M. Blight, J. A. Pickett, and B. J. Pye. 1994. Development of field strategies incorporating
semiochemicals for the control of pea and bean weevil, Sitona lineatus L. Crop Prot. 13:127-135.
Hanavan, R. P., N. A. Bosque-Perez, D. J. Schotzko, S. O. Guy, and S. D. Eigenbrode. 2008. Early season aerial adult
colonization and ground activity of pea leaf weevil (Coleoptera: Curculionidae) in pea as influenced by tillage system. J.
Econ. Entomol. 101:1606-1613.
Hanavan, R. P., N. A. Bosque-Perez, D. J. Schotzko, and S. D. Eigenbrode. 2010. Influence of tillage on adult and
immature pea leaf weevil (Coleoptera: Curculionidae) densities in pea. J. Econ. Entomol. 103:691-697.
Hatten, T. D., R. M. Dahlquist, S. D. Eigenbrode, and N. A. Bosque-Perez. 2010. Tillage affects the activity-density,
absolute density, and feeding damage of the pea leaf weevil in spring pea. Entomol. Exp. Appl. 136:235-242.
Eilenberg, J., A. Hajek, and C. Lomer. 2001. Suggestions for unifying the terminology in biological control. Biocontrol
46:387-400. 30.
R. J. R005. Host specificity testing and suitability of a
European biotype of the braconid parasitoid Microctonus aethiopoides as a biological control agent against Sitona lepidus
(Coleoptera: Curculionidae) in New Zealand Biocont. Sci. Technol. 15:791-813.
De Clerk-Floate, R., and H. Cárcamo. 2011. Biocontrol arthropods: new denizens of Canada's grassland agro-ecosystems.
In: K. Floate (ed.) Vol. II – Arthropods of Canadian Grasslands (in press).
Aeschlimann, J.-P. 1977. Notes on Patasson lameerei (Hym.: Mymaridae) an egg parasitoid of Sitona spp (Col.:
Curculionidae) in the Mediterranean region. Entomophaga 22:111-114.
Jaworska, M. 1998. Laboratory preference of annual legumes by pea weevil Sitona lineatus L. (Col. Curculionidae) and
their effect on susceptibility of weevils to entomogenous nematodes. J. Invertebr. Pathol. 71:248-250.
Hamon, N., R. Bardner, L. Allen-Williams, and J. B. Lee. 1990. Carabid populations in field beans and their effect on the
Sitona lineatus (L.) Ann. Appl. Biol. 117:51-62.
Vankosky, M. A., H. A. Cárcamo, and L. M. Dosdall. 2010. Identification of potential natural enemies of the pea leaf
Sitona lineatus L. in western Canada. J. Appl. Entomol. doi: 10.1111/j.1439-0418.2010.01542.x.
Floate, K. D., H. A. Cárcamo, R. E. Blackshaw, B. Postman, and S. Bourassa. 2007. Response of ground beetle
(Coleoptera: Carabidae) field populations to four years of Lepidoptera-specific Bt corn production. Environ. Entomol.
36:1269-1274. 37.
Ripper, W. E., R. M. Greenslade, and L. A. Lickerish. 1949. Combined chemical and biological control of insects by
means of a systemic insecticide. Nature 163:787-789.
Maienfisch, P., M. Angst, F. Brandl, W. Fischer, D. Hofer, H. Kayser, W. Kobel, A. Rindlisbacher, R. Senn, A.
Steinemann, and H. Widmer. 2001. Chemistry and biology of thiamethoxam: a second generation neonicotinoid. Pest
Manage. Sci. 57:906-913.
Elbert, A., M. Haas, B. Springer, W. Thielert, and R. Nauen. 2008. Applied aspects of neonicotinoid uses in crop
protection. Pest Manage. Sci. 64:1099-1105.
Volume 4 ▪ 2011
www.prairiesoilsandcrops.ca
Source: http://prairiesoilsandcrops.ca/articles/volume-4-9-screen.pdf
karplab.net
Nanocarriers as an emerging platform for cancer therapy Nanotechnology has the potential to revolutionize cancer diagnosis and therapy. Advances in protein engineering and materials science have contributed to novel nanoscale targeting approaches that may bring new hope to cancer patients. Several therapeutic nanocarriers have been approved for clinical use. However, to date, there are only a few clinically approved nanocarriers that incorporate
Protocolo actuación urgencias sanitarias en los centros de castilla y león
PROTOCOLOS DE ACTUACIÓN ANTEURGENCIAS SANITARIAS EN LOS CENTROSEDUCATIVOS DE CASTILLA Y LEÓN Actualmente, la promoción de la salud en los centros docentes es una de las metas prioritarias que noshemos marcado los responsables educativos de la Junta de Castilla y León, pues consideramos que promoverla salud entre los escolares, en sus múltiples aspectos o vertientes, significa favorecer su desarrollo personal,su bienestar físico, su equilibrio emocional y, por tanto, su calidad de vida.